— 生态 · 农业 · 健康 —

时间:2020-02-17
作者:易科泰
点击量:
简介:
FKM多光谱荧光动态显微成像系统

FKM(Fluorescence Kinetic Microscope)多光谱荧光动态显微成像系统是目前功能最为强大全面的植物显微荧光研究仪器,是基于FluorCam叶绿素荧光成像技术的显微成像定制系统。它由包含可扩展部件的增强显微镜、高分辨率CCD相机、激发光源组、光谱仪、控温模块以及相应的控制单元和专用的工作站与分析软件组成。它不仅可以进行微藻、单个细胞、单个叶绿体乃至基粒-基质类囊体片段进行Fv/Fm、Kautsky诱导效应、荧光淬灭、OJIP快速荧光响应曲线、QA再氧化等各种叶绿素荧光及MCF多光谱荧光(multicolor fluorescence)成像分析;还能通过激发光源组进行进行任意荧光激发和荧光释放波段的测量,从而进行GFP、DAPI、DiBAC4、SYTOX、CTC等荧光蛋白、荧光染料以及藻青蛋白、藻红蛋白、藻胆素等藻类特有荧光色素的成像分析;更可以利用光谱仪对各种荧光进行光谱分析,区分各发色团(例如PSI和PSII及各种捕光色素复合体等)并进行深入分析。
FKM多光谱荧光动态显微成像系统使荧光成像技术真正成为光合作用机理研究的探针,使科研工作者在藻类和高等植物细胞与亚细胞层次深入理解光合作用过程及该过程中发生的各种变化,为直接研究叶绿体中光合系统的工作机理提供了最为有力的工具。FKM作为藻类/植物表型和基因型显微研究的双重利器,得到了学界的广泛认可并取得了大量的科研成果。
功能特点

应用领域

测量样品
工作原理
FKM分析过程中,通过连接在显微镜上的激发光源组和内置在6位滤波轮中的一系列滤波器、分光镜激发植物样品中各种发色团的动态荧光。样品激发出的荧光经显微镜放大后进行荧光光谱分析和荧光动力学成像分析。SM 9000光谱仪通过光纤与显微镜连接,以进行激发荧光光谱分析。安装在显微镜顶部的高分辨率CCD相机则用于荧光动力学成像分析。全部工作过程通过工作站和控制单元按照预先设定好的程序自动进行。测量过程中,可通过温控模块调控藻类、植物细胞等实验样品的温度。蠕动泵可以实现培养藻类的连续测量。
仪器组成
1 增强显微镜
2 高分辨率CCD相机
3 激发光源组
4 SM 9000光谱仪
5 主控制单元
6 工作站及软件
7 控温模块的控制单元
8 位滤波轮

技术参数
测量参数
具备完备的自动测量程序(protocol),可自由对自动测量程序进行编辑
荧光激发光源:
透射光源(选配):白光、远红光
高分辨率TOMI-2 CCD传感器:
显微镜:Axio Imager M2,可选配Axio Scope A1简洁版或Axio Imager Z2高级版
物镜:10倍、20倍、40倍、63倍和100倍专用生物荧光物镜(可选)
6位滤波轮:叶绿素荧光、GFP/SYTOX、DAPI/CTC等
SM9000光谱仪
温度调控模块:温度调节范围为环境温度(室内温度)至70℃,精确度0.1℃
蠕动泵(选配):流速10-5600µl/min,用于藻类连续培养测量
FluorCam叶绿素荧光成像分析软件功能:具Live(实况测试)、Protocols(实验程序选择定制)、Pre–processing(成像预处理)、Result(成像分析结果)等功能菜单
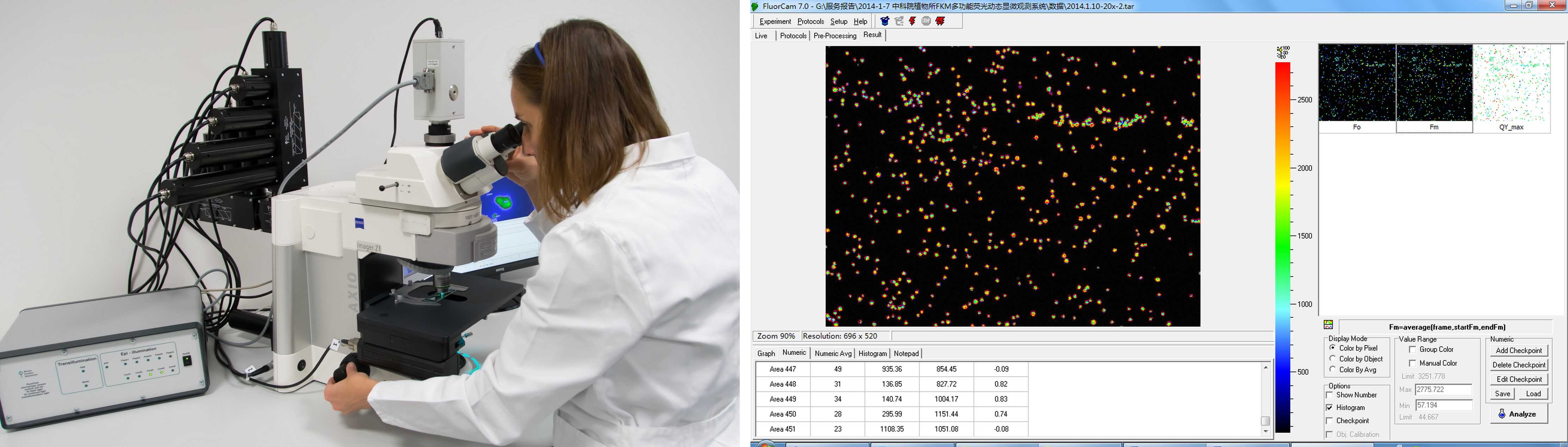
叶绿素荧光与光谱分析结果
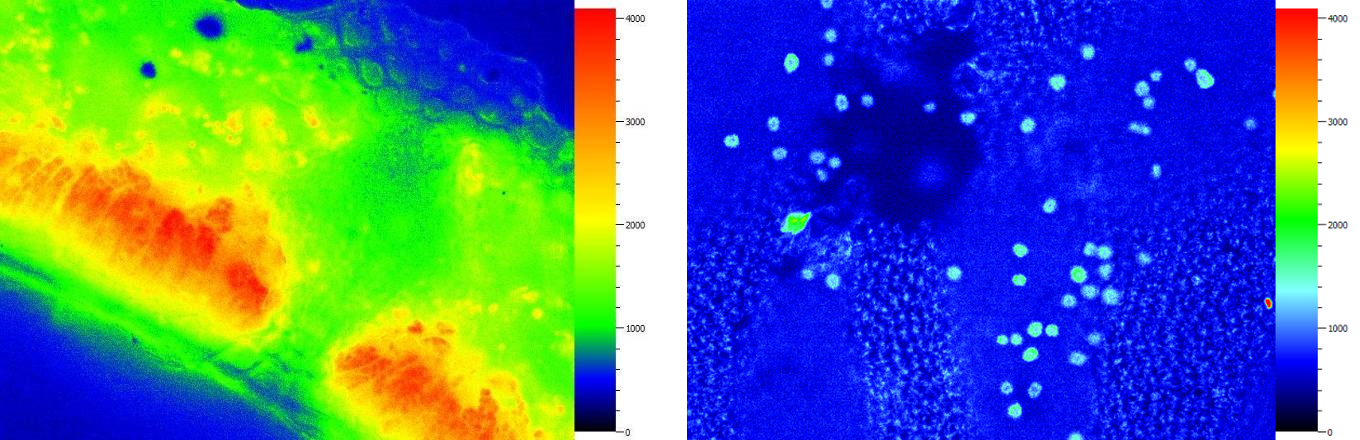
忍冬叶片横切荧光成像——栅栏组织和海绵组织 硅藻附生的紫菜表面荧光成像——细胞内的叶绿体分布

鸢尾表皮细胞荧光成像——气孔与叶绿体 衣藻荧光成像
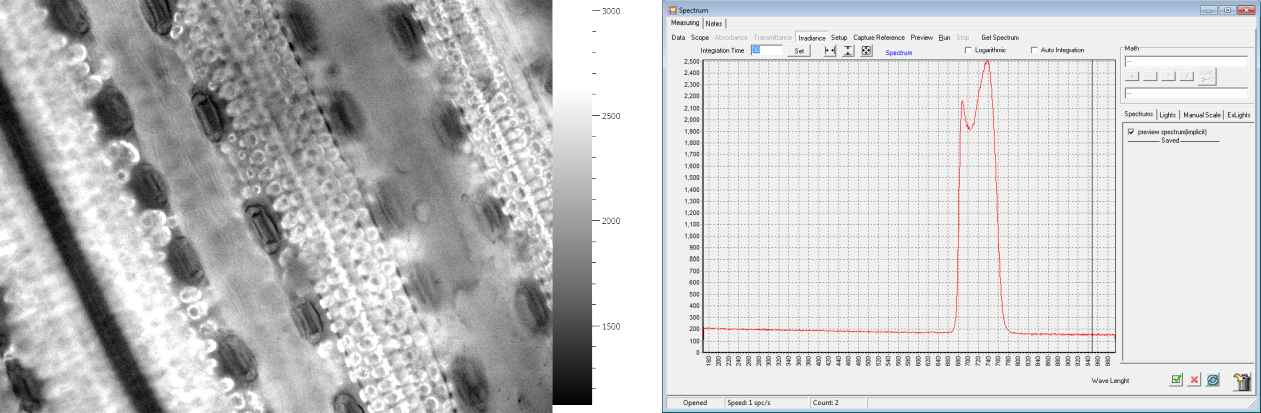
玉米表皮细胞荧光成像 叶绿素荧光光谱分析
典型应用
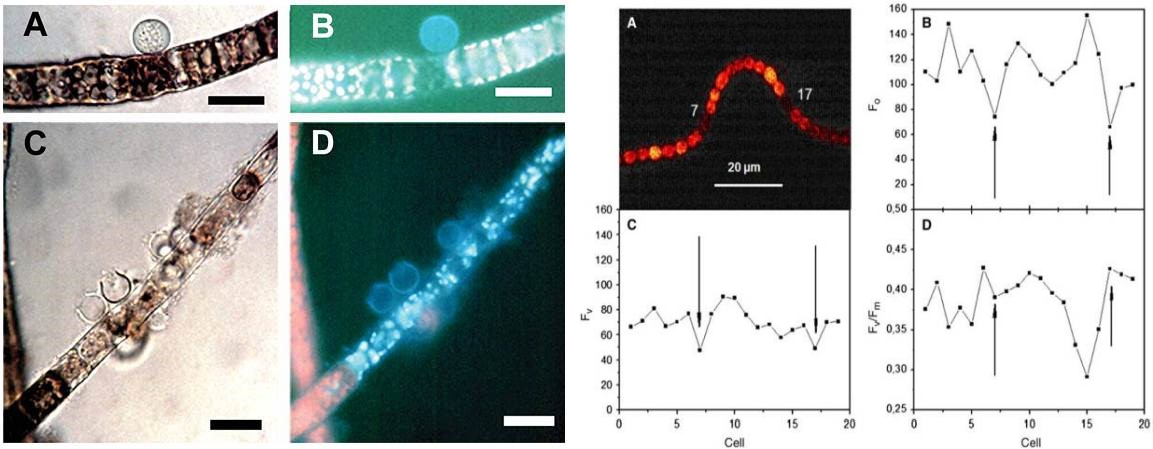
藻类病害(Gachon C, et al. 2006) 藻类异形胞光合生理与变化过程 (Ferimazova N, et al. 2013)
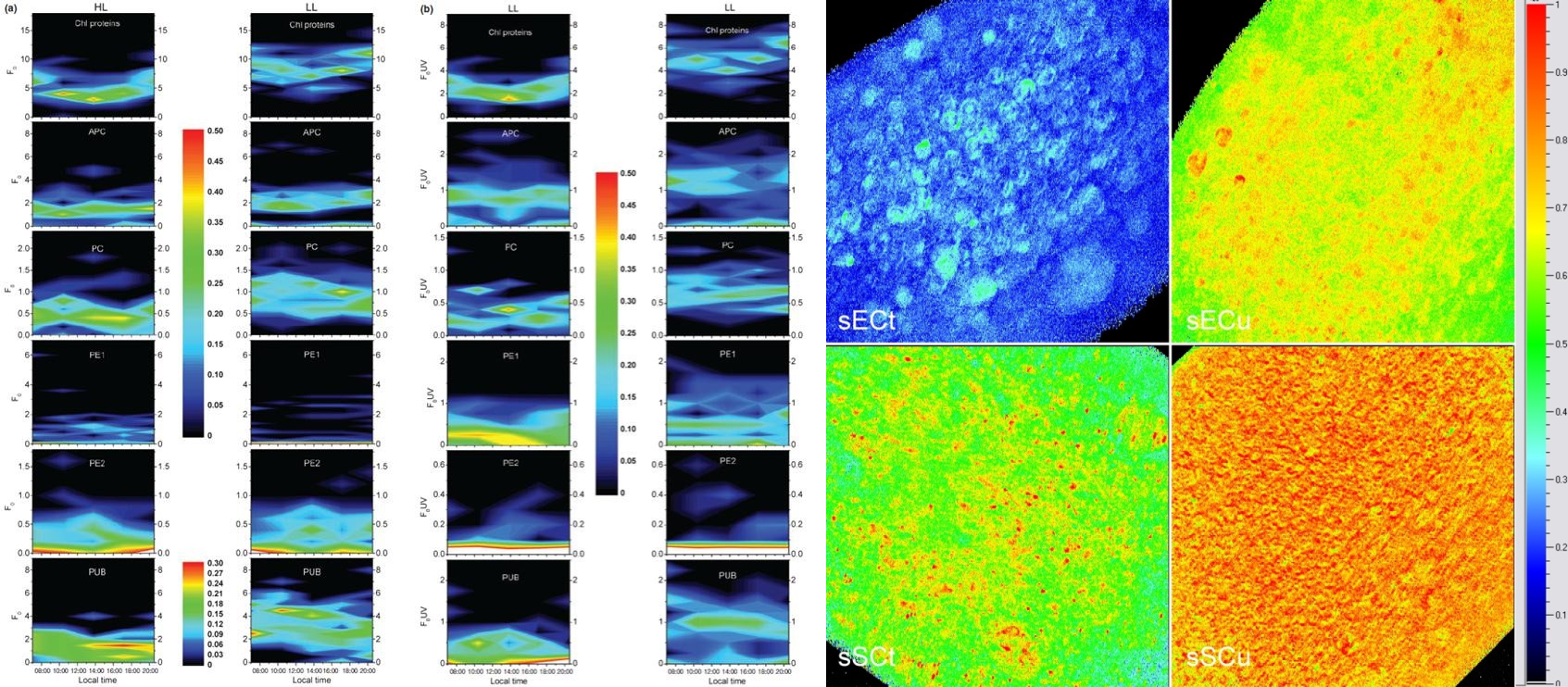
藻类特有不同光合色素蛋白复合体的荧光成像与分析(Andresen, et al. 2010) 重金属胁迫对藻类/植物的影响(Thomas G, et al. 2016)
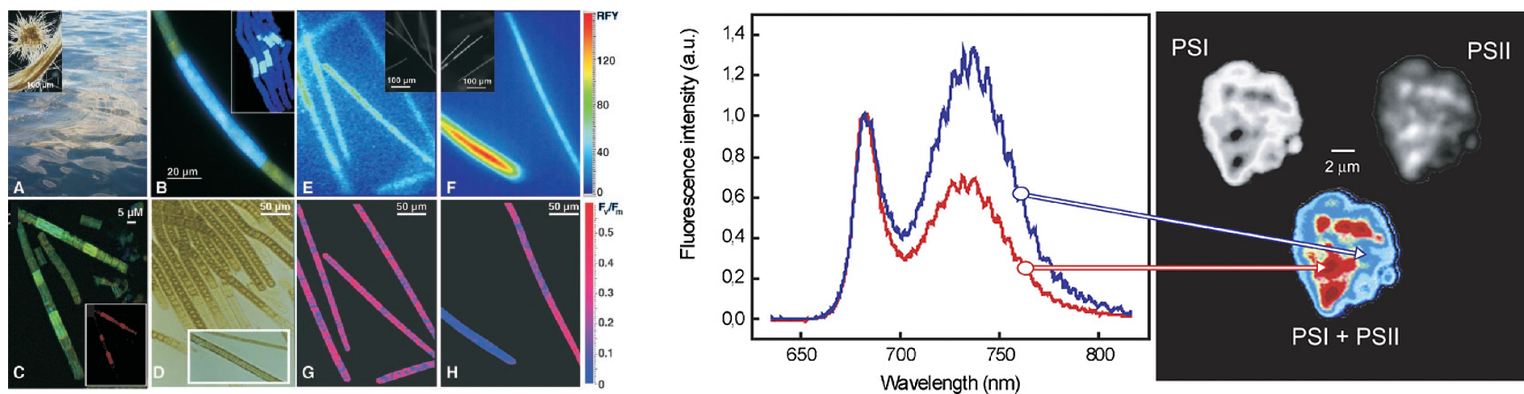
蓝藻光合与固氮的时空隔离(Berman-Frank I, et al. 2001) 植物/藻类光合作用机制的深入研究(Vacha F, et al. 2007.)
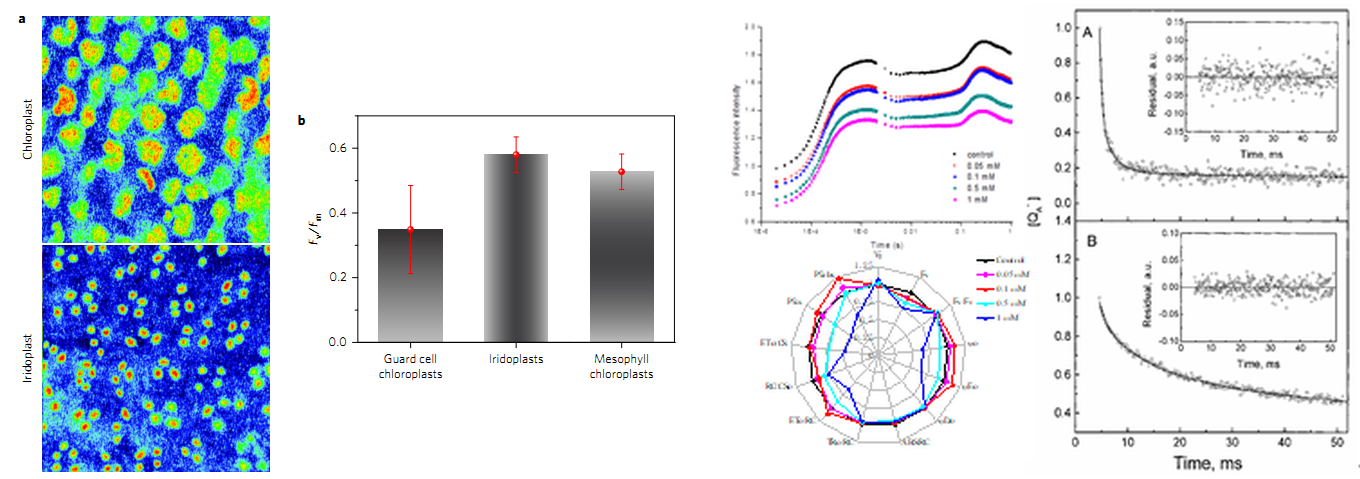
单个细胞\叶绿体荧光动力学与表型分析(Jacobs M, et al. 2016.) OJIP快速荧光曲线和QA再氧化动力学曲线测量
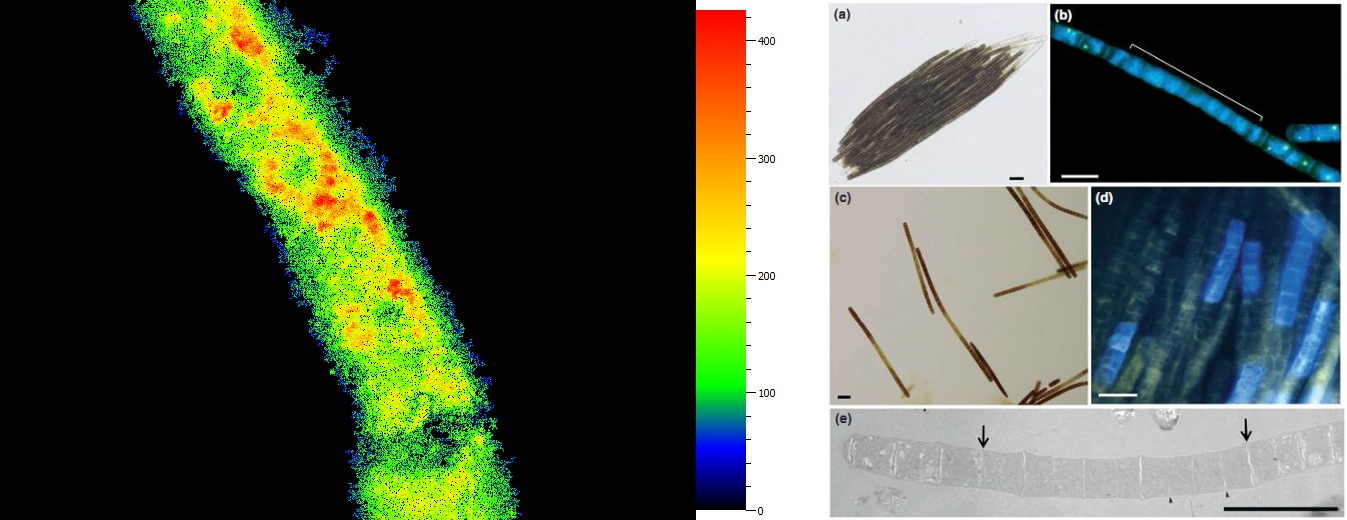
C4植物玉米花环结构光合特性分析 荧光蛋白与荧光染料显微成像
产地:捷克
参考文献:
1)Morina F, et al. 2021. Interactions between zinc and Phomopsis longicolla infection in the roots of Glycine max. Journal of Experimental Botany 72(8):3320-3336
2)Ashraf N, et al. 2021. Effect of nanomolar concentrations of lanthanum on Desmodesmus quadricauda cultivated under environmentally relevant conditions. Aquatic Toxicology 235: 105818
3)Umeki M, et al. 2020. Fecal pellets of giant clams as a route for transporting Symbiodiniaceae to corals. PLoS ONE 15(12): e0243087
4)Morina F, & Küpper H. 2020. Direct inhibition of photosynthesis by Cd dominates over inhibition caused by micronutrient deficiency in the Cd/Zn hyperaccumulator Arabidopsis halleri. Plant Physiology and Biochemistry. DOI:10.1016/j.plaphy.2020.07.018
5)Morina F, et al. 2020. Interaction Between Zn Deficiency, Toxicity and Turnip Yellow Mosaic Virus Infection in Noccaea ochroleucum. Frontiers in Plant Science, 11.DOI:10.3389/fpls.2020.00739
6)Wang ZX, et al. 2020. Extravagant leaves display in Actinidia kolomikta attracting pollinators and maintaining photosynthetic capacity. bioRxiv, DOI: 10.1101/2020.03.05.979500
7)刘文娟等,2020. 两个玉米品种维管束鞘叶绿体的非光化学淬灭对干旱胁迫的响应. 中国农业科学 53(8): 1532-1544
8)Andresen E, et al. 2019. Chronic exposure of soybean plants to nanomolar cadmium reveals specific additional high-affinity targets of Cd toxicity. Journal of Experimental Botany. DOI:10.1093/jxb/erz530
9)Morishima SY, et al. 2019. Study on expelled but viable zooxanthellae from giant clams, with an emphasis on their potential as subsequent symbiont sources. PLOS ONE, 14(7), e0220141.
10)Kupper H, et al. 2018. Analysis of OJIP chlorophyll fluorescence kinetics and QA re-oxidation kinetics by direct fast imaging. Plant Physiology, pp.00953.2018.

西安研发中心
微信公众号

业务咨询
微信号

易科泰公司
微信公众号

易科泰公司
视频号
联系我们:
地址: 北京市海淀区高里掌路3号院6号楼1单元101B
电话: 010-82611269/1572
手机: 13671083121
传真: 010-62465844
Email: info@eco-tech.com.cn
友情链接:

